skip to main |
skip to sidebar


Las proteínas son macromoléculas formadas por cadenas lineales de aminoácidos. El nombre proteína proviene de la palabra griega πρώτα ("prota"), que significa "lo primero" o del dios proteo, por la cantidad de formas que pueden tomar. Características
Las proteínas son moléculas de enorme tamaño; pertenecen a la categoría de macromoléculas; son polímeros, es decir, están constituidas por gran número de unidades estructurales simples repetitivas (monómeros). Debido a su gran tamaño, cuando estas moléculas se dispersan en un disolvente adecuado, forman siempre dispersiones coloidales, con características que las distinguen de las soluciones de moléculas más pequeñas.
Por hidrólisis, las moléculas proteínicas son escindidas en numerosos compuestos relativamente simples, de pequeño peso, que son las unidades fundamentales constituyentes de la macromolécula. Estas unidades son los aminoácidos, de los cuales existen veinte especies diferentes y que se unen entre sí mediante enlaces peptídicos. Cientos y miles de estos aminoácidos pueden participar en la formación de la gran molécula polimérica de una proteína.
Todas las proteínas contienen carbono, hidrógeno, oxígeno y nitrógeno y casi todas poseen también azufre. Si bien hay ligeras variaciones en diferentes proteínas, el contenido de nitrógeno representa, término medio, 16% de la masa total de la molécula; es decir, cada 6,25 g de proteínas contienen 1 g de N. El factor 6,25 se utiliza para estimar la cantidad de proteína existente en una muestra a partir de la medición de N de la misma.
La síntesis proteica es un proceso complejo cumplido por las células según las directrices de la información suministrada por los genes.
Funciones
Las proteínas ocupan un lugar de máxima importancia entre las moléculas constituyentes de los seres vivos (biomoléculas). Prácticamente todos los procesos biológicos dependen de la presencia y/o actividad de este tipo de sustancias. Bastan algunos ejemplos para dar idea de la variedad y trascendencia de funciones a ellas asignadas. Son proteínas casi todas las enzimas, catalizadores de reacciones químicas en organismos vivientes; muchas hormonas, reguladores de actividades celulares; la hemoglobina y otras moléculas con funciones de transporte en la sangre; los anticuerpos, encargados de acciones de defensa natural contra infecciones o agentes extraños; los receptores de las células, a los cuales se fijan moléculas capaces de desencadenar una respuesta determinada; la actina y la miosina, responsables finales del acortamiento del músculo durante la contracción; el colágeno, integrante de fibras altamente resistentes en tejidos de sostén

Introducción
Los carbohidratos, hidratos de carbono y también simplemente azúcares. En su composición entran los elementos carbono, hidrógeno y oxígeno, con frecuencia en la proporción Cn(H20)n, por ejemplo, glucosa C6(H2O)6 de aquí los nombres carbohidratos o hidratos de carbono.
Estos compuestos, abarcan sustancias muy conocidas y al mismo tiempo, bastante disímiles, azúcar común, papel, madera, algodón, son carbohidratos o están presentes en ello en una alta proporción.
A partir del dióxido de carbono y agua, las plantas sintetizan los carbohidratos, en un proceso denominado fotosíntesis.
El pigmento verde de las plantas, la clorofila, pone a disposición del vegetal, la energía que absorbe de la luz solar. En este proceso tienen lugar numerosas reacciones catalizadas por enzimas, no todas se comprenden, queda el CO2 reducido como carbohidrato y a su vez se libera oxígeno.
La energía solar quedó transformada en energía química a disposición de las plantas y de animales, los cuales metabolizan los carbohidratos realizando la operación inversa y utilizando la energía para diversos fines.
Ingerimos cereales, pero los cereales, digamos arroz, maíz, contienen almidones, estos son macromoléculas poliméricas de glucosa, que nuestro organismo procesa y transforma con sus enzimas para nuestro beneficio:
La glucosa, no solamente la utiliza el organismo como fuente de energía, puede transformarla en otras macromoléculas, el glucógeno, que se acumula en el hígado y músculos y sirve de reserva de energía, la transforma en colesterol y hormonas esferoidales imprescindibles para numerosas funciones. Si se ingieren excesos de carbohidratos estos se transforman en grasas. De modo que, estos compuestos resultan importantes para nosotros, no solo por el algodón, papel y madera, los carbohidratos constituyen uno de los tres grandes grupos de alimentos.
Clasificación Los carbohidratos se clasifican en monosacáridos, oligosacáridos y polisacáridos. Un monosacárido, es una unidad, ya no se subdivide más por hidrólisis ácida o enzimática, por ejemplo glucosa, fructosa o galactosa.
Los oligosacáridos están constituidos por dos a diez unidades de monosacáridos. La palabra viene del griego, oligo = pocos. Digamos el azúcar que utilizamos es un disacárido y por tanto un oligosacárido.
Los polisacáridos son macromoléculas, por hidrólisis producen muchos monosacáridos, entre 100 y 90 000 unidades.
Como primera aproximación, desde el punto de vista químico, los carbohidratos son polihidroxialdehídos o polihidroxicetonas o compuestos que los producen por hidrólisis ácida o enzimática. Esto es solo parcialmente cierto, pues en solución acuosa, las estructuras de polihidroxialdehídos o de polihidroxicetonas, permanecen en pequeña proporción en equilibrio con sus formas cíclicas, que son las más abundantes. Estos aspectos interesantes los veremos más adelante.Monosacáridos
Como ya señalamos, en una primera aproximación, son polihidroxialdehídos o polihidroxicetonas. La estructura contiene pues, varios grupos hidroxilos y un grupo carbonilo. El sufijo que se utiliza al referirnos a ellos es "osa". Una hexosa es por tanto, un monosacárido de seis átomos de carbono. Si el carbonilo se presenta como aldehído será una aldohexosa y si se presenta de forma similar a una cetona, diremos es una cetohexosa.
La mayoría de los monosacáridos naturales son pentosas o hexosas.
Pentosa Hexosa Hexosa
Aldopentosa Aldohexosa Cetohexosa
Para representar estructuras de carbohidratos, se utiliza una representación abreviada, las fórmulas de proyección de Fischer. Las fórmulas de proyección de Fischer, resultan cómodas para representar estructuras y por tanto, se continúan utilizando, igual que el convenio de clasificar los carbohidratos como pertenecientes a las familias D o L, en lugar de utilizar el convenio mucho más actual de clasificar R o S (Cahn-Igold-Prelog). Digamos D(+) gliceraldehido, D porque el –OH está a la derecha y el signo (+) se refiere solo a la rotación de luz polarizada, es una molécula dextrógira. Así un carbohidrato que presenta el –OH del estereocentro más alejado del carbonilo a la derecha, se clasifica como D. si estuviera a la izquierda, se clasifica como perteneciente a la familia L o serie L.
Algunas aldopentosas naturales:
D-Ribosa 2- Dexoxi- D-Xilosa D-Arabinosa
D-ribosa
La ribosa y la dexoxiribosa forman parte de los ácidos nucleicos. La ribosa también se aisla de la hidrólisis de la riboflavina (vitamina B2). El prefijo "dexoxi" se refiere a que este monosacárido contiene menos átomos de oxígeno que lo común, incumple con la fórmula Cn(H20)n.
La xilosa y la arabinosa, pueden aislarse de los productos de hidrólisis de las resinas vegetales, recibiendo la xilosa también la denominación de "azúcar de madera". La D(-) Arabinosa se encuentra también en bacterias y esponjas. Las hexosas naturales más comunes son:
D(+)- Glucosa D(+)-Manosa D(+)-Galactosa L(+)- Ramnosa D(-)- Fructosa
La glucosa también recibe el nombre de dextrosa por ser dextrorrotatoria (D(+)-Glucosa), también azúcar de sangre, pues está presente en la sangre humana en concentración de 65-110 mg/100 ml. Es posiblemente el producto natural más abundante pues se encuentra como polisacárido en el almidón, la celulosa y el glucógeno. También aparece combinada como disacárido en el azúcar común, la sacarosa (fructosa y glucosa) y en la leche de todos los mamíferos, lactosa, azúcar de leche (galactosa y glucosa).
La glucosa, galactosa y ramnosa forman con frecuencia parte de glicósidos naturales. Los glicósodos son compuestos con una estrucura formada por uno o más carbohidratos que se enlazan a una molécula que no es un carbohidrato. El conjunto se llama glicósido y la porción que no es un carbohidrato se denomina aglicón.
La fructosa es un ejemplo de cetohexosa, es entre los azúcares el compuesto más dulce, tiene bastante más poder edulcorante que la sacarosa, donde se encuentra enlazada con la glucosa. Esta cetohexosa se encuentra libre en la miel y en muchas frutas.
La D(+)-Manosa, se encuentra formando muchos polisacáridos naturales.
.
Propiedades químicas de los monosacáridos
Muchas reacciones de los monosacáridos, son debidas a la pequeña cantidad de forma abierta, acíclica, en equilibrio con las estructuras cíclicas. Algunas reacciones que requieren una concentración inicial mayor fallan, el fallo se debe a que la forma aldehídica no tiene concentración suficiente para que se produzca la reacción, no obstante los monosacáridos presentan una variedad de reacciones que se producen bien, las reacciones son la típicas de las funciones presentes, carbonilo e hidrixilo y por supuestos a interacciones entre ambos grupos.
Reacciones de los grupos hidroxilos
Acilación.
Los grupos hidroxilos de los monosacáridos, se pueden esterificar. Por ejemplo tratando la glucosa u otra hexosa con anhídrido acético, se pueden obtener los derivados pentacetilados estereoisoméricos, el α o el β. El curso estereoquνmico de la reacciσn, depende de los catalizadores y otras condiciones.
Para ver la fórmula seleccione la opción "Descargar" del menú superior
α, β-glucopiranosa α, β-pentacetilglucopiranosa
3.3. Estereoquímica de los monosacáridos
En ocasiones has encontrado términos tales como, dextrógira o levógira, la designación están referida a las propiedades que presentan los carbohidratos en disolución de originar un giro en el plano de vibración de la luz polarizada, bien hacia la derecha o hacia la izquierda.
En el primer caso se denominan dextrorrotatorias y en el segundo levorrortatorias. Estos compuestos que presentan esta propiedad se plantea que son óptimamente activos y el fenómeno se conoce como actividad óptica, (isómeros ópticos, estereoisómeros).
La luz polarizada.
La luz polarizada es un fenómeno electromagnético. Un rayo de luz consiste en dos campos oscilantes mutuamente perpendiculares: un campo eléctrico y un campo magnético.
Si se pudiera observar un rayo de luz ordinaria en un extremo, pudiéramos ver que los planos en los cuales la oscilación eléctrica se produce, están en todas direcciones y perpendiculares a la dirección de la propagación. Cuando la luz ordinaria pasa a trabes de un polarizador, el polarizador interactúa con el campo eléctrico de forma tal que en el campo eléctrico de la luz que emerge del polarizador (y el campo magnético asociado a él) está oscilando en un plano. Tal luz es llamada luz polarizada en un plano.
Luz ordinaria Luz polarizada en un plano
El polarímetro es el equipo que se utiliza para determinar actividad óptica.
Fuente de luz Polarizador Tubo con la muestra Analizador
Si el analizador rota en el sentido de las agujas del reloj se dice que es (+) y si lo hace en sentido contrario es negativa (-).
(+) (-)
dextrorrotatoria levororrotatoria
El número de estereoisómeros de una de las estructuras de los monosacáridos, se puede calcular utilizando la fórmula del químico alemán Emil Fischer:
N=2n
N= número de estereoisómeros
n= números de carbonos estereogénicos (estereocentros)
Por lo tanto para las aldohexosas tendremos N=24=16 estereoisómeros.
Átomo de carbono con cuatro átomos o grupos de átomos diferentes. Se señala con un asterisco (*).
Enantiómeros.
Existen estereoisómeros cuya relación entre sí, es el de objeto-imagen. Estos estereoisómeros se conocen como Enantiómeros, el fenómeno recibe el nombre de enantiomería.
Diasteroisómeros.
Parejas de estereoisómeros que no son imágenes especulares.
Es importante darse cuenta de que los conceptos de Enantiómeros y Diasteroisómeros carecen de sentido considerados aisladamente.
Para ver la fórmula seleccione la opción "Descargar" del menú superior
D (+)- Glucosa D (+)-Manosa
(I) (II) (III)
I y II Enantiómeros.
II y III Diasteroisómeros.
Oligosacáridos
Generalidades sobre los disacáridos.
Como su nombre lo indica un disacárido, es un carbohidrato formado por dos unidades de monosacáridos. Estas unidades están unidas mediante un enlace glicosídico.
¿Qué es un enlace glicosídico?
Es un éter formado en el hidroxilo hemiacetálico y se clasifica bajo la denominación de Glicósido.
Para ver la fórmula seleccione la opción "Descargar" del menú superior
Un carbono hemiacetálico, es aquel al que están unidos, un hidrógeno, un hidroxilo, un grupo R-O- y un grupo R.
Un glicósido sera:
Donde R" puede ser un carbohidrato u otro tipo de molécula que no es un carbohidrato (la porción que no es azúcar se le llama aglicón).
En los disacáridos existe unión glicosídica, entre dos monosacáridos. En ellos es importante saber: cómo se verifica esta unión, a dónde, en qué posición está situado el enlace.
Una unión muy común es la 1-4´, quiere decir, están comprometidos en el enlace, el C1, carbono anomérico de una unidad y el C4 de la otra unidad.
β
Para ver el gráfico seleccione la opción "Descargar" del menú superior
Enlace 1-4´ β
En este tipo de unión queda libre el hidroxilo del carbono anomérico de la segunda unidad y por lo tanto, el disacárido en solución acuosa estará en equilibrio con la forma aldehídica. También ocurre lo mismo, en el otro tipo muy común de unión, la 1-6´.
Disacárido con enlaceβ 1-4´
Un disacárido con enlace 1-4´ ó 1-6´, está capacitado para establecer equilibrio con la forma aldehídica y por tanto, presentará las reacciones típicas de los aldehídos que ya estudiamos; reducirá los reactivos de Tollens, Fehling y Benedict. Por el contrario, si el enlace compromete los dos carbonos anoméricos, no hay posibilidades de que se establezca el equilibrio con la forma aldehídica y por tanto este disacárido no presentará ninguna de las propiedades y reacciones que hemos mencionado.
El enlace entre los monosacáridos se define como α σ β en dependencia de la estereoquνmica del enlace de la primera unidad.
Como sabemos, todos los monosacáridos son reductores, la mayoría de los disacáridos también.
Cuatro disacáridos naturales más importantes son la sacarosa, la celobiosa, la lactosa y la maltosa.
Celobiosa.
La celobiosa se obtiene por hidrólisis parcial de la celulosa, es un disacárido formado por dos unidades de glucosas unidas por enlaces β-1,4´.
Para ver el gráfico seleccione la opción "Descargar" del menú superior
Sacarosa.
El azúcar de mesa, sacarosa, es un disacárido formado por fructosa y glucosa, es el disacárido más abundante en el reino vegetal.
La sacarosa no es reductora, pues el enlace entre los dos monosacáridos está formado por los hidroxilos de los carbonos anoméricos.
Para ver el gráfico seleccione la opción "Descargar" del menú superior
Sacarosa: α-glucopiranosil-β- D- fructofuranσsido
Lactosa.
Es un disacárido formado por una unidad de galactosa y otra de glucosa, unidas por enlaces β-1,4΄.
La lactosa, está presente en la leche de los mamíferos, la leche de vaca contiene de 4-6 % y la humana de 5-8 %.
La lactosa puede también existir en dos formas puede ser α, o puede ser β. A la temperatura del cuerpo la leche materna consiste aproximadamente en una mezcla de 2 partes de α-lactosa y tres de β-lactosa.
Maltosa.
La maltosa, es un alimento para niños. Es un disacárido formado por dos unidades de glucosa con enlace α-1,4΄.
4-O-(α-glucopiranosil)-D-glucopiranosa
De este disacárido se conocen las dos formas en dependencia de la estereoquímica del hidroxilo hemiacetálico que puede ser α, o puede ser β que es la forma habitual de la maltosa.
Polisacáridos.
Son polímeros naturales, macromoléculas, formadas por monosacáridos, cientos de unidades enlazadas y a veces están constituidas por miles de unidades. Dos ejemplos típicos de polisacáridos son el almidón y la celulosa.
Almidón.
Reserva energética de las plantas y para nosotros un alimento. Se encuentra en forma en forma de pequeños granos en muchas partes, u órganos constituyentes de las plantas, especialmente en semillas y tejidos vegetales embrionarios, en tubérculos de papa, semillas de arroz, maíz o trigo. Ellos sirven de nutrientes para el proceso germinativo y en general para el desarrollo de las plantas.
Como primera aproximación, se puede decir que el almidón está constituido por unidades de D(+)-glucosa enlazadas α-1,4´. Nuestras enzimas hidrolizan los almidones hasta sus unidades constituyentes de glucosa, la cual, como ya hemos expresado, sirve a nuestro organismo de nutriente y es utilizada para diferentes transformaciones metabólicas.
Al tratar el almidón con agua caliente, este se separa en dos fracciones: una dispersable, que se conoce como amilasa y otra no dispersable, que es la mayoritaria, que se conoce como amilopectina.
Para ver el gráfico seleccione la opción "Descargar" del menú superior
Los dos constituyentes del almidón difieren en diversos aspectos y por tanto los consideramos por separado.
Amilosa
Para ver el gráfico seleccione la opción "Descargar" del menú superior
Amilopectina.
Está también constituido por unidades de D(+)-glucosa, con enlaces α-1,4΄, pero las cadenas son de 1000 unidades o mαs y presentan ramificaciones cada 25 unidades α-1,6΄.
La hidrólisis de la amilopectina van produciendo mezclas de Oligosacáridos, de masa moleculares gradualmente menores, que se conocen como "Dextrinas" que se utilizan en el acabado de tejidos y en la fabricación de pegamentos, etc.
Para ver el gráfico seleccione la opción "Descargar" del menú superior
Amilopectina
Por hidrólisis parcial, se puede detectar la presencia de isomaltosa, disacárido que tiene enlaces α-1,6΄,
Para ver el gráfico seleccione la opción "Descargar" del menú superior
Celulosa.
La celulosa es el polisacárido más abundante en la naturaleza, es el tejido de sostén de las plantas, formando aproximadamente la mitad de las paredes o membranas de las células vegetales. Pero la celulosa, no está sola, está asociada con las hemicelulas y la lignina. La celulosa está formada por unidades de D(+)-glucosa, los enlaces en el polisacáridos son β 1,4´: este tipo de enlace los carnívoros no pueden romperlo y por tanto no pueden utilizar la glucosa como nutriente.
Para ver el gráfico seleccione la opción "Descargar" del menú superior
Celulosa
La celulosa forma microfibrillas de 14 000 unidades o más, que se torcionan y se unen a otras por puente de hidrógeno. La masa molecular varía entre 250 000 y 1 x 106 o más.
Si se trata celulosa con NaOH con concentración de 17,5 %, una parte (cadenas menores) resulta soluble, la β celulosa. Lo que no resulta disuelto (cadenas mayores) la llaman α celulosa.
Para obtener pulpa de celulosa, con un 90 % de pureza a partir de madera, existen diversos procedimientos: en uno calientan virutas de madera con solución de hidróxido de sodio, o bien con solución de hidrógenosulfito de sodio y ácido sulfuroso; usan digestores, reactores a presión a 4 atm. En este procedimiento de "celulosa de sulfito", las hemicelulosas resultan disueltas y también las ligninas (en forma de sulfonatos). Este "licor de sulfito" lo utilizan para la producción colateral de etanol. La pulpa de celulosa, lavada, la tratan con "polvos de blanqueo" (mezclas de clorato de calcios, cloruro e hidróxido de calcio).
Para la producción del papel de filtro y papel de alta calidad, se aplican posteriores purificaciones y en el caso del papel, se utilizan aditivos como el sulfato de bario o sulfato de calcio y colofonia para cerrar los poros.
Derivados de la celulosa.
Durante décadas el hombre, en su esfuerzo por vivir mejor ha desarrollado derivados de la celulosa, por ejemplo el nitrato de celulosa, con contenido de nitrógeno mayor a 11 %, está constituidos por mezclas de di y trinitrato, se utiliza como explosivo de impulsión. Esto se prepara a partir del algodón y con mezcla nitrante (HNO3/H2SO4), por lo que se conoce como "algodón pólvora" y también pólvora sin humo.
El acetato de celulosa: los tres hidroxilos de la glucosa de la celulosa, pueden ser acetilados. Para ello se trata la celulosa con una mezcla de ácido acético y anhídrido acético, adicionando cantidad catalítica de H2SO4 (conc.). Este triacetato, se hridroliza obteniéndose diacetato, cadenas de 200 a 300 unidades, con esto se obtienen películas de fotografías. También a partir del diacetato, se obtienen hilos, fibra textil, láminas para retroproyector, películas, etc.
Los ésteres metílicos y etílicos, de la celulosa que son plásticos de importancia comercial.

Los lípidos son biomoléculas orgánicas formadas básicamente por carbono e hidrógeno y generalmente también oxígeno; pero en porcentajes mucho más bajos. Además pueden contener también fósforo, nitrógeno y azufre .
Es un grupo de sustancias muy heterogéneas que sólo tienen en común estas dos características:
Son insolubles en agua
Son solubles en disolventes orgánicos, como éter, cloroformo, benceno, etc.
Una característica básica de los lípidos, y de la que derivan sus principales propiedades biológicas es la hidrofobicidad. La baja solubilidad de los lipídos se debe a que su estructura química es fundamentalmente hidrocarbonada (alifática, alicíclica o aromática), con gran cantidad de enlaces C-H y C-C (Figura de la izquierda). La naturaleza de estos enlaces es 100% covalente y su momento dipolar es mínimo. El agua, al ser una molécula muy polar, con gran facilidad para formar puentes de hidrógeno, no es capaz de interaccionar con estas moléculas. En presencia de moléculas lipídicas, el agua adopta en torno a ellas una estructura muy ordenada que maximiza las interacciones entre las propias moléculas de agua, forzando a la molécula hidrofóbica al interior de una estructura en forma de jaula, que también reduce la movilidad del lípido. Todo ello supone una configuración de baja entropía, que resulta energéticamente desfavorable. Esta disminución de entropía es mínima si las moléculas lipídicas se agregan entre sí, e interaccionan mediante fuerzas de corto alcance, como las fuerzas de Van der Waals. Este fenómeno recibe el nombre de efecto hidrofóbico (Figuras inferiores).
Para ver el gráfico seleccione la opción "Descargar" del menú superior
Constituyentes importantes de la alimentación (aceites, manteca, yema de huevo), representan una importante fuente de energía y de almacenamiento, funcionan como aislantes térmicos, componentes estructurales de membranas biológicas, son precursores de hormonas (sexuales, corticales), ácidos biliares, vitaminas etc.
FUNCIONES DE LOS LÍPIDOS
Los lípidos desempeñan cuatro tipos de funciones:
Función de reserva. Son la principal reserva energética del organismo. Un gramo de grasa produce 9'4 kilocalorías en las reacciones metabólicas de oxidación, mientras que proteínas y glúcidos sólo producen 4'1 kilocaloría/gr.
Función estructural. Forman las bicapas lipídicas de las membranas. Recubren órganos y le dan consistencia, o protegen mecánicamente como el tejido adiposo de piés y manos.
Función biocatalizadora. En este papel los lípidos favorecen o facilitan las reacciones químicas que se producen en los seres vivos. Cumplen esta función las vitaminas lipídicas, las hormonas esteroideas y las prostaglandinas.
Función transportadora. El tranporte de lípidos desde el intestino hasta su lugar de destino se raliza mediante su emulsión gracias a los ácidos biliares y a los proteolípidos.
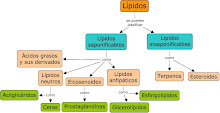
CLASIFICACIÓN DE LOS LÍPIDO Los lípidos se clasifican en dos grupos, atendiendo a que posean en su composición ácidos grasos (Lípidos saponificables) o no lo posean ( Lípidos insaponificables ).
1. Lípidos saponificables
A. Simples
Acilglicéridos
Céridos
B. Complejos
Fosfolípidos
Glucolípidos
2. Lípidos insaponificables
A. Terpenos
B. Esteroides
C. Prostaglandinas
ÁCIDOS GRASOS
Los ácidos grasos son moléculas formadas por una larga cadena hidrocarbonada de tipo lineal, y con un número par de átomos de carbono. Tienen en un extremo de la cadena un grupo carboxilo (-COOH).
Para ver el gráfico seleccione la opción "Descargar" del menú superior
Se conocen unos 70 ácidos grasos que se pueden clasificar en dos grupos :
Los ácidos grasos saturados sólo tienen enlaces simples entre los átomos de carbono. Son ejemplos de este tipo de ácidos el mirístico (14C);el palmítico (16C) y el esteárico (18C) .
Los ácidos grasos insaturados tienen uno o varios enlaces dobles en su cadena y sus moléculas presentan codos, con cambios de dirección en los lugares dónde aparece un doble enlace. Son ejemplos el oléico (18C, un doble enlace) y el linoleíco (18C y dos dobles enlaces).
Para ver el gráfico seleccione la opción "Descargar" del menú superior
Propiedades de los ácidos grasos
Solubilidad. Los ácidos grasos poseen una zona hidrófila, el grupo carboxilo (-COOH) y una zona lipófila, la cadena hidrocarbonada que presenta grupos metileno (-CH2-) y grupos metilo (-CH3) terminales. Por eso las moléculas de los ácidos grasos son anfipáticas, pues por una parte, la cadena alifática es apolar y por tanto, soluble en disolventes orgánicos (lipófila), y por otra, el grupo carboxilo es polar y soluble en agua (hidrófilo).
Desde el punto de vista químico, los ácidos grasos son capaces de formar enlaces éster con los grupos alcohol de otras moléculas. Cuando estos enlaces se hidrolizan con un álcali, se rompen y se obtienen las sales de los ácidos grasos correspondientes, denominados jabones, mediante un proceso denominado saponificación.
Para ver el gráfico seleccione la opción "Descargar" del menú superior
LÍPIDOS SIMPLESSon lípidos saponificables en cuya composición química sólo intervienen carbono, hidrógeno y oxígeno.
Acilglicéridos Son lípidos simples formados por la esterificación de una,dos o tres moléculas de ácidos grasos con una molécula de glicerina. También reciben el nombre de glicéridos o grasas simples
Para ver el gráfico seleccione la opción "Descargar" del menú superior
Según el número de ácidos grasos, se distinguen tres tipos de estos lípidos:
los monoglicéridos, que contienen una molécula de ácido graso
los diglicéridos, con dos moléculas de ácidos grasos
los triglicéridos, con tres moléculas de ácidos grasos.
Los acilglicéridos frente a bases dan lugar a reacciones de saponificación en la que se producen moléculas de jabón.
Ceras Las ceras son ésteres de ácidos grasos de cadena larga, con alcoholes también de cadena larga. En general son sólidas y totalmente insolubles en agua. Todas las funciones que realizan están relacionadas con su impermeabilidad al agua y con su consistencia firme. Así las plumas, el pelo , la piel,las hojas, frutos, están cubiertas de una capa cérea protectora.
Una de las ceras más conocidas es la que segregan las abejas para confeccionar su panal.
LÍPIDOS COMPLEJOS
Son lípidos saponificables en cuya estructura molecular además de carbono, hidrógeno y oxígeno, hay también nitrógeno,fósforo, azufre o un glúcido. Son las principales moléculas constitutivas de la doble capa lipídica de la membrana, por lo que también se llaman lípidos de membrana. Son tammbién moléculas anfipáticas.
Fosfolípidos Se caracterizan pr presentar un ácido ortofosfórico en su zona polar. Son las moléculas más abundantes de la membrana citoplasmática.
Algunos ejemplos de fosfolípidos
Para ver los gráficos seleccione la opción "Descargar" del menú superior
Glucolípidos Son lípidos complejos que se caracterizan por poseer un glúcido. Se encuentran formando parte de las bicapas lipídicas de las membranas de todas las células, especialmente de las neuronas. Se sitúan en la cara externa de la membrana celular, en donde realizan una función de relación celular, siendo receptores de moléculas externas que darán lugar a respuestas celulares.
Terpenos
Son moléculas lineales o cíclicas que cumplen funciones muy variadas, entre los que se pueden citar:
Esencias vegetales como el mentol, el geraniol, limoneno, alcanfor, eucaliptol,vainillina.
Vitaminas, como la vit.A, vit. E, vit.K.
Pigmentos vegetales, como la carotina y la xantofila.
Esteroides
Los esteroides son lípidos que derivan del esterano. Comprenden dos grandes grupos de sustancias:
Esteroles: Como el colesterol y las vitaminas D.
Hormonas esteroideas: Como las hormonas suprarrenales y las hormonas sexuales.
Para ver el gráfico seleccione la opción "Descargar" del menú superior
El colesterol forma parte estructural de las membranas a las que confiere estabilidad. Es la molécula base que sirve para la síntesis de casi todos los esteroides
HORMONAS SEXUALES
Entre las hormonas sexuales se encuentran la progesterona que prepara los órganos sexuales femeninos para la gestación y la testosterona responsable de los caracteres sexuales masculinos.
Para ver el gráfico seleccione la opción "Descargar" del menú superior
HORMONAS SUPRARRENALES
Entre las hormonas suprarrenales se encuentra la cortisona, que actúa en el metabolismo de los glúcidos, regulando
regulando la síntesis de glucógeno.
Para ver el gráfico seleccione la opción "Descargar" del menú superior
Prostaglandinas
Las prostaglandinas son lípidos cuya molécula básica está constituída por 20 átomos de carbono que forman un anillo ciclopentano y dos cadenas alifáticas.
Para ver el gráfico seleccione la opción "Descargar" del menú superior
Las funciones son diversas. Entre ellas destaca la producción de sustancias que regulan la coagulación de la sangre y cierre de las heridas; la aparición de la fiebre como defensa de las infecciones; la reducción de la secreción de jugos gástricos. Funcionan como hormonas locales.
BIBLIOGRAFÍA
Bioquímica de Harper
Química – Chang
Bioquímica - Horton, H. Robert; Moran, Laurence A; Ochs Raymond S; Rawn, J. David; Scrimgeour K. Gray - México, D.F: Prentice-Hall Hispanoamericana, 1995
Química - Sienko, Michell J; Plane, Robert A - Madrid: Aguilar, 1967

Colesterol
Síntesis del colesterol
El colesterol es un lípido que se encuentra en los tejidos corporales y en el plasma sanguíneo de los vertebrados. Se presenta en altas concentraciones en el hígado, médula espinal, páncreas y cerebro. El nombre de «colesterol» procede del griego kole (bilis) y stereos (sólido), por haberse identificado por primera vez en los cálculos de la vesícula biliar por Michel Eugène Chevreul quien le dio el nombre de «colesterina».
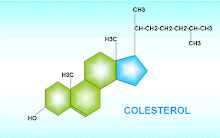
Estructura química
Es un lípido esteroide, molécula de ciclopentanoperhidrofenantreno (o ester), constituida por cuatro carbociclos condensados o fundidos, denominados A, B, C y D, que presentan varias sustituciones:
Dos radicales metilo en las posiciones C-10 y C-13.
Una cadena no metálica en la posición C-17.
Un grupo hidroxilo en la posición C-3.
Una insaturación entre los carbonos C-5 y C-6.
En la molécula de colesterol se puede distinguir una cabeza polar constituida por el grupo hidroxilo y una cola o porción apolar formada por el carbociclo de núcleos condensados y los sustituyentes alifáticos. Así, el colesterol es una molécula tan hidrófoba que la solubilidad de colesterol libre en agua es de 10-8 M y, al igual que los otros lípidos, es bastante soluble en disolventes apolares como el cloroformo (CCl4).
Fuentes de colesterol
Los organismos mamíferos obtienen colesterol a través de dos vías:
1. Vía exógena o absorción de colesterol pre-existente en los alimentos. El colesterol se encuentra exclusivamente en los alimentos de origen animal, mayoritariamente la yema de huevo, hígado, lácteos, cerebro (sesos) y músculo esquelético (carnes rojas).
2. Vía endógena o síntesis de novo, es la síntesis de colesterol en las células animales a partir de su precursor, el acetato, en su forma activada acetil-coenzima A
Síntesis de colesterol
La biosíntesis del colesterol tiene lugar en el retículo endoplásmico (liso) de virtualmente todas las células de los animales vertebrados. Mediante estudios de marcaje isotópico, D. Rittenberg y K. Bloch demostraron que todos los átomos de carbono del colesterol proceden, en última instancia, del acetato, en forma de acetil-Coenzima A. Se requirieron aproximadamente otros 30 años de investigación para describir las líneas generales de la biosíntesis del colesterol, desconociéndose, sin embargo, muchos detalles enzimáticos y mecanísticos a la fecha. Los pasos principales de la síntesis de colesterol son:
El acetil-CoA se convierte en mevalonato.
El mevalonato se convierte en escualeno mediante reacciones sucesivas de transferencia de grupos prenilo.
El escualeno se transforma en lanosterol.
El lanosterol se convierte en colesterol después de otras 21 reacciones sucesivas, enzimáticamente catalizadas.
Transporte del colesterol
Debido a su gran insolubilidad en agua el colesterol circula en la sangre exclusivamente asociado a complejos macromoleculares conocidos como lipoproteínas.
Regulación del colesterol
La producción de colesterol es regulada directamente por la concentración del colesterol presente en el retículo endoplásmico de las células, habiendo una relación indirecta con los niveles plasmáticos de colesterol presente en las lipoproteínas de baja densidad (LDL por su acrónimo inglés). Una alta ingesta de colesterol en los alimentos conduce a una disminución neta de la producción endógena y viceversa. El principal mecanismo regulador de la homeostasis de colesterol celular aparentemente reside en un complejo sistema molecular centrado en las proteínas SREBPs (Sterol Regulatory Element Binding Proteins 1 y 2: proteínas que se unen a elementos reguladores de esteroles). En presencia de una concentración crítica de colesterol en la membrana del retículo endoplásmico, las SREBPs establecen complejos con otras dos importantes proteínas reguladoras: SCAP (SREBP-cleavage activating protein: proteína activadora a través del clivaje de SREBP) e Insig (insulin induced gene) 1 y 2. Cuando disminuye la concentración del colesterol en el retículo endoplásmico, las Insigs se disocian del complejo SREBP-SCAP, permitiendo que el complejo migre al aparato de Golgi, donde SREBP es escindido secuencialmente por S1P y S2P (site 1 and 2 proteases: proteasas del sitio 1 y 2 respectivamente). El SREBP escindido migra al núcleo celular donde actúa como factor de transcripción uniéndose al SRE (Sterol Regulatory Element: elemento regulador de esteroles) de una serie de genes relevantes en la homeostasis celular y corporal de esteroles, regulando su transcripción. Entre los genes regulados por el sistema Insig-SCAP-SREBP destacan los del receptor de lipoproteínas de baja densidad (LDLR) y la hidroxi-metil-glutaril CoA-reductasa (HMG-CoA-reductasa), la enzima limitante en la vía biosintética del colesterol.
Tras dilucidar los mecanismos celulares de captación endocítica de colesterol lipoproteico, trabajo por el cual fueron galardonados con el premio Nobel en fisiología y medicina en el año 1985, Michael S. Brown y Joseph L. Goldstein han participado directamente en el descubrimiento y caracterización de la vía de los SREBPs de regulación del colesterol corporal. Estos avances han sido la base del mejor entendimiento de la fisiopatología de diversas enfermedades humanas, fundamentalmente la enfermedad vascular aterosclerótica, principal causa de muerte en el mundo occidental a través del infarto agudo al miocárdio y los accidentes cerebrovasculares y el fundamento de la farmacología de las drogas hipocolesteromiantes más potentes: las estatinas.
Funciones del colesterol
El colesterol es imprescindible para la vida por sus numerosas funciones:
Estructural: el colesterol es un componente muy importante de las membranas plasmáticas de los animales (no existe en los vegetales). Aunque el colesterol se encuentra en pequeña cantidad en las membranas celulares, en la membrana citoplasmática lo hallamos en una proporción molar 1:1 con relación a los fosfolípidos, regulando sus propiedades físico-químicas, en particular la fluidez. Sin embargo, el colesterol se encuentra en muy baja proporción o está prácticamente ausente en las membranas subcelulares.
Precursor de la vitamina D: esencial en el metabolismo del calcio.
Precursor de las hormonas sexuales: progesterona, estrógenos y testosterona.
Precursor de las hormonas corticoesteroidales: cortisol y aldosterona.
Precursor de las sales biliares: esenciales en la absorción de algunos nutrientes lipídicos y vía principal para la excreción de colesterol corporal.
Precursor de las balsas de lípidos...










{kind=link}